
肿瘤的发生发展不仅取决于基因的突变或缺失,还随着肿瘤细胞的代谢重塑或异常改变而发生改变。在营养缺乏的条件下,肿瘤细胞的代谢重编程赋予癌细胞快速增殖的能力。其中,氨基酸代谢重编程是肿瘤代谢异常改变的重要特征之一。研究发现,氨基酸不仅能够作为氮供体为肿瘤细胞的增殖、侵袭和免疫逃逸过程提供核苷酸等生物大分子的合成原料,而且还是肿瘤微环境中免疫细胞活化和发挥抗肿瘤作用的重要代谢物质。氨基酸代谢的异常改变与肿瘤的发生发展和肿瘤免疫密切相关,其代谢途径中的部分关键蛋白质或关键酶可作为肿瘤诊断和治疗的生物标志物。因此,本文围绕氨基酸转运体对癌细胞增殖的影响和肿瘤代谢循环过程中的谷氨酰胺、天冬酰胺、丝氨酸和甘氨酸等氨基酸代谢的异常改变进行总结,介绍了氨基酸代谢与肿瘤细胞mTOR 信号通路、肿瘤微环境和免疫细胞功能的相关性,对靶向氨基酸代谢的肿瘤治疗药物进行了分析和展望。期望该工作为深入了解氨基酸代谢对肿瘤发生发展的调控及其可能存在的肿瘤治疗靶点提供有用的参考。
上个世纪30 年代末,研究人员首次发现肿瘤细胞具有瓦博格效应(Warburg effect)。该发现不仅引领了从代谢的角度研究肿瘤发生发展的新方向[1]
,还揭示了肿瘤细胞内部的葡萄糖代谢重编程现象。随着对肿瘤代谢逐步深入的研究,发现肿瘤细胞为满足其增殖所需的能量要求,通常出现能量和物质代谢重编程的现象[2] 。然而,值得注意的是,某些肿瘤细胞在增殖或转移过程中存在不依赖葡萄糖而更依赖谷氨酰胺的情况[3] 。研究表明,除了糖代谢能够为肿瘤细胞增殖提供能量外,氨基酸代谢也在肿瘤的发生发展过程中具有不可替代的作用。
在肿瘤的发生发展过程中,癌细胞内氨基酸的摄取速率、氨基酸的代谢途径、代谢产物或代谢关键酶出现异常的现象称之为氨基酸代谢重编程。其中,肿瘤细胞内的氨基酸除了直接参与蛋白质合成外,还通过分解代谢产物与其他对肿瘤发生至关重要的代谢途径相关联。在许多人类癌症中,均发现氨基酸代谢重编程与肿瘤发展具有相关性,包括卵巢癌、肝细胞癌、肾透明细胞癌和乳腺癌等[4-8] 。另外,研究发现,精氨酸、色氨酸等对于T 细胞的活化和分化具有重要功能[9] ;同样,肿瘤微环境(tumormicro-environment, TME)中的某些氨基酸的缺乏或氨基酸代谢物的产生,对于免疫细胞的功能维持也很重要。这表明,氨基酸不仅在肿瘤细胞增殖过程中能够提供能量,还能通过调控免疫细胞功能间接参与肿瘤免疫过程。因此,了解肿瘤的氨基酸代谢重编程及其参与肿瘤免疫的机制,对于研究肿瘤异常代谢的发生发展及寻找肿瘤的治疗靶点具有重要指导意义。
1 与肿瘤代谢相关的氨基酸转运体
肿瘤细胞在增殖过程中,为满足其能量需求,需要大量摄取氨基酸,但是由于氨基酸具有亲水性,不能以自由扩散的形式被肿瘤细胞摄取和利用,因此,需要各种氨基酸转运体的协助。这意味着肿瘤细胞的增殖与氨基酸转运体的表达密切相关。为满足癌细胞摄取氨基酸的需求,氨基酸转运体(heteromericamino acid transports, HATs)能够转运肿瘤细胞所需的多种必需氨基酸,并参与调控肿瘤细胞摄取氨基酸的速率和维持胞内氨基酸的水平[10] 。而且有研究发现,多种氨基酸转运体能够促进癌细胞中雷帕霉素靶蛋白( mammalian target of rapamycin,mTOR)的活化和肿瘤细胞增殖。因此,氨基酸转运体对于肿瘤细胞的发生发展具有调控功能,通过靶向氨基酸转运体实现对肿瘤生长的抑制是一个有效的策略[11] 。
1. 1 谷氨酰胺转运体
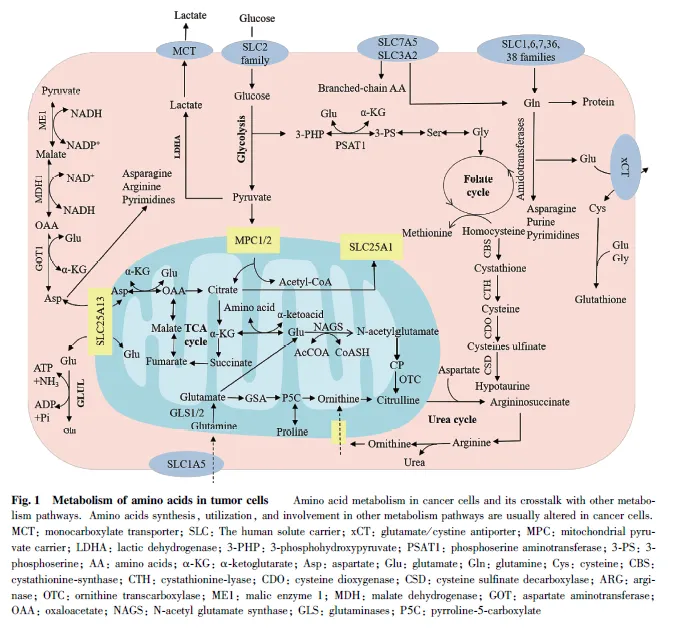
在正常细胞内,丙氨酸,丝氨酸,半胱氨酸转运蛋白2 ( alanine, serine, cysteine transporter 2,ASCT2)属于溶质载体家族1 (solute carrier family1, SLC1)成员,负责谷氨酰胺和天冬酰胺的转运。然而,研究发现,ASCT2 在多种类型的癌细胞中表达量上调,并帮助肿瘤细胞摄取更多的谷氨酰胺用于增殖[12] 。而肿瘤细胞的快速增殖又进一步促进了ASCT2 的表达,由此形成了肿瘤细胞异常增殖的恶性循环。在多种异种移植模型中发现, 敲低ASCT2 会降低mTORC1 的活性并抑制肿瘤的生长[13] 。除此之外,钠离子依赖的中性氨基酸转运蛋白2(sodium-dependent neutral amino acid transporter2, SNAT2)也是谷氨酰胺的转运体之一,然而,值得注意的是,谷氨酰胺主要通过ASCT2 转运进入癌细胞[14] ,当肿瘤中ASCT2 缺失时,细胞会上调SNAT2的表达从而替代ASCT2 的功能。若同时缺失ASCT2 和SNAT2 则会减缓肿瘤细胞的生长[15] 。研究表明,SNAT2 对于肿瘤细胞的增殖同样具有调控作用。
1. 2 必需氨基酸转运体
必需氨基酸对于癌细胞的增殖是必不可少的,它们通过细胞膜的摄取受到膜转运蛋白质的严格控制,因此,必需氨基酸转运体对调控肿瘤细胞的增殖具有重要作用。其中,L-型氨基酸转运蛋白1(Ltypeamino acid transporter 1, LAT1)能转运肿瘤细胞所需的中性氨基酸。在许多类型的癌症中都发现LAT1 高表达,且LAT1 上调可导致人类肿瘤细胞功能异常[16] 。研究发现,在结直肠癌细胞中,由于基因的突变会导致谷氨酰胺的摄取增多,从而改变了细胞的代谢。在此过程中,LAT1 会在特异性基因激活后维持细胞氨基酸水平。因此,其对早期和晚期转移性疾病模型中的结直肠肿瘤发生至关重要[17] 。并且LAT1 是调节肿瘤血管内皮生成和肿瘤增殖的关键分子[18] ,抑制LAT1 可导致某些类型的癌症发生细胞凋亡。例如,T 细胞急性淋巴细胞白血病、胆管癌和骨肉瘤等[19-21] 。其他研究也表明,在人类巨噬细胞、B 细胞和自然杀伤细胞中,敲除LAT1 或抑制其活性会降低亮氨酸摄取、mTOR 复合物1(mTOR complex 1, mTORC1)活性、糖酵解速率和c-Myc 水平,并且进一步降低白介素-1β(Interleukin-1β, IL-1β)的产生[22-23] 。研究表明,LAT1不仅与肿瘤细胞内的信号通路相互作用参与肿瘤细胞增殖调控,还通过调节免疫细胞生长或细胞因子的产生,参与肿瘤免疫过程。
1. 3 胱氨酸/ 谷氨酸反向转运蛋白
胱氨酸/ 谷氨酸反向转运蛋白(system Xc-)是氨基酸转运蛋白质家族的成员,由轻链xCT 和重链4F2hc 这2 个亚基组成。在正常细胞中,溶质载体家族7 成员11(solute carrier family 7 member 11,SLC7A11)属于胱氨酸/ 谷氨酸逆向转运蛋白,其能将氧化的半胱氨酸转运至细胞,并将胞内产生的谷氨酸排出细胞[24] (见Fig. 1)。SLC7A11 能够通过促进谷胱甘肽的生物合成,来降低氧化应激反应,从而参与细胞增殖调控。肿瘤细胞由于代谢途径的异常,对抗氧化机制具有很强的依赖性,已有研究表明,阻断SLC7A11/ 谷胱甘肽轴选择性地抑制鼠类肉瘤病毒癌基因(kirsten rat sarcoma viral oncogene,Kras)突变的非小细胞肺癌的生长[25-26] 。因此,谷胱甘肽的平衡和抗氧化能力的增强对于肿瘤细胞的增殖具有重要意义。有研究证实,xCT 在多种类型的肿瘤中出现高表达,并与肿瘤增殖、侵袭、转移和耐药性有关[27] ,并且ASCT2 和xCT 在氧化还原和代谢方面具有协同效应,因此,癌细胞中xCT 和ASCT2的基因缺失会导致肿瘤生长减慢,这使得它们成为抗癌药物开发的靶点[28] 。
值得注意的是,SLC7A11 是铁死亡的抑制因子[29] ,能够促进癌细胞的存活。因此,转录因子NF-E2 相关因子2(nuclear erythroid 2-related
factor2, Nrf2)和p53 为发挥肿瘤调控功能,通常协同参与调控SLC7A11 的表达来发挥抗氧化功能,并影响肿瘤细胞铁死亡进程[30-31] 。与之类似的是,在肺癌细胞中RNA 结合基元单链结合蛋白1(RNA bindingmotif, single
stranded interacting protein 1, RBMS1)可以通过调控SLC7A11 的翻译来影响肿瘤细胞的铁死亡,从而介导肺癌细胞的生长调控[32] 。研究表明,氨基酸转运体SLC7A11 不仅能负责氨基酸的转运,还能调控肿瘤细胞的铁代谢。另一方面,与SLC7A11介导肿瘤细胞铁死亡机制不同的是,在葡萄糖缺乏的情况下,SLC7A11能介导通过磷酸戊糖途径产生的烟酰胺腺嘌呤二核苷酸(nicotinamideadeninedinucleotide
phosphate, NADPH) 将胱氨酸还原为半胱氨酸。如果肿瘤细胞在高表达SLC7A1情况下,缺少葡萄糖,则会导致细胞发生氧化应激后快速死亡。这进一步表明,SLC7A11 介导的胱氨酸代谢与癌细胞的其他代谢之间存在关联,其能够协调磷酸戊糖途径的进程来参与肿瘤细胞代谢重编程[33] 。
2 肿瘤代谢循环过程中的氨基酸
2. 1 谷氨酰胺及谷氨酸代谢
谷氨酰胺是被机体摄取最多的氨基酸, 参与一系列的生物反应,包括能量生成、大分子合成以及信号转导。在多种癌细胞中,谷氨酰胺是仅次于葡萄糖为肿瘤细胞提供能量的物质,因此,与肿瘤的发生发展密切相关。由于癌细胞的氨基酸需求增加,致使谷氨酰胺在细胞快速增殖过程中始终依赖于外源供应,即肿瘤细胞的“谷氨酰胺依赖现象”[34] 。肿瘤细胞的增殖会消耗氧气,导致肿瘤微环境处于缺氧状态,因此,为满足其能量要求其主要碳源来源于谷氨酰胺[35] 。研究发现,在免疫细胞、小肠细胞和一些癌细胞在内的快速增殖细胞中,谷氨酰胺除了提供氮、碳的生物合成反应外,还在细胞能量学和降低癌细胞氧化应激方面发挥重要作用[36] 。由于谷氨酰胺酶(glutaminase, GLS)介导谷氨酰胺的分解代谢,并能够为肿瘤增殖提供所需原料,目前,关于GLS 调控肿瘤生长的机制研究较多。例如,胰腺导管癌的生长和存活对于谷氨酰胺表现出很强的依赖性,其细胞GLS 的表达水平较高。且高表达的GLS通常出现琥珀酰化修饰来增强自身活性,这样能通过增强谷氨酰胺的分解代谢来促进细胞的存活[37] 。而且,肿瘤细胞增殖过程中的大多数非必需氨基酸都直接来源于谷氨酰胺[38] 。
研究发现,在肿瘤细胞的谷氨酰胺合成过程中,谷氨酰胺代谢酶受到肿瘤细胞转录因子的精确调控,这些转录因子可以通过影响谷氨酰胺代谢酶(glutamine synthetase, GS)的表达水平和酶活性来调控谷氨酰胺代谢和肿瘤细胞的生物学活动。例如,转录因子c-Myc 可以抑制miR23a/ b 的表达来回补三羧酸(TCA)循环,并在此过程中上调GLS 的表达,从而上调谷氨酸代谢量[39]。与之类似的是,肿瘤抑制因子p53 也能通过与苹果酸酶(malic en-zyme, ME)基因的共有反应元件结合来抑制所有ME 同种型基因的表达,从而调节谷氨酰胺代谢、NAPDH 产生、氧化还原状态以及生物合成[40] 。有趣的是,除了与转录因子相互作用参与肿瘤代谢调控外,最新发现,GS 能够通过直接促进肿瘤细胞的有丝分裂来增强其增殖,而这种调控细胞周期的功能却独立于GS 的代谢酶功能[41] 。以上研究表明,谷氨酰胺在细胞增殖、生物合成反应和氧化还原稳态提供碳和还原氮中具有重要功能。另外,在肿瘤细胞发展过程中,消耗血清中的谷氨酰胺、阻断细胞对谷氨酰胺的摄取,以及抑制参与谷氨酰胺合成或分解代谢的酶等均能影响肿瘤细胞的增殖。因此,靶向谷氨酰胺代谢过程中的部分关键酶和中间代谢物,已被证实可以调控肿瘤代谢过程,甚至进一步影响肿瘤的生长。
2. 2 天冬氨酸和天冬酰胺代谢
天冬氨酸与天冬酰胺属于非必需氨基酸,可部分依赖于体内生物合成。在肿瘤细胞内,天冬氨酸和天冬酰胺不仅会参与肿瘤细胞的增殖,还会调控癌细胞内的信号传导。研究发现,天冬酰胺合成酶(asparagine synthetase, ASNS)在体内的表达量密切影响着癌症的发生发展,肿瘤细胞在谷氨酰胺不足或缺失的情况下,外源补充天冬酰胺可维持蛋白质的翻译并促进肿瘤细胞的生长[42] 。与之类似的是,在缺乏谷氨酰胺的内皮细胞中,天冬酰胺对于恢复蛋白质合成、抑制内质网应激以及重新激活mTOR信号传导至关重要[43] 。进一步的研究发现,敲除天冬酰胺合成酶或限制饮食中的天冬酰胺,可减少肿瘤的转移而不影响原发性肿瘤的生长,而增加饮食中的天冬酰胺或增强天冬酰胺合成酶表达会促进肿瘤转移发生[44] 。之前的研究已发现,ASNS 的表达缺陷会导致细胞对天冬酰胺的限制异常敏感,通过实验进一步证实,限制天冬酰胺可激活肿瘤细胞内的其他信号通路,限制与此有关的信号通路,将会使得肿瘤细胞的生长更容易因天冬酰胺的限制受到限制[45] 。值得注意的是,除了直接调控肿瘤的转移外,天冬酰胺还能通过与肿瘤内其他信号分子相互作用以间接调控肿瘤的发展。有研究发现,p53 通过转录下调ASNS 表达抑制天冬酰胺合成,并且能通过破坏天冬酰胺-天冬氨酸稳态,抑制体内和体外淋巴瘤和结肠肿瘤生长[46] 。这些研究表明,不仅外源的天冬酰胺会影响肿瘤细胞的增殖,内源天冬酰胺水平的稳定对于肿瘤细胞内部信号传导也具有调控作用。
2. 3 丝氨酸和甘氨酸代谢
丝氨酸和甘氨酸对于肿瘤细胞的增殖极其重要,其代谢过程中的部分代谢物或关键酶会调控肿瘤细胞的生长。当丝氨酸外源补充不足时,肿瘤细胞可通过丝氨酸合成途径(serine synthesis pathway,SSP)产生内源性丝氨酸。在癌细胞中,糖酵解的产物三磷酸甘油酸,在磷酸甘油酸脱氢酶(phosphoglyceratedehydrogenase,
PHGDH)和NADPH 的参与下生成磷酸羟丙酮酸,再经过一系列反应生成丝氨酸。PHGDH 作为丝氨酸合成的关键酶,在调控肿瘤细胞增殖和迁移过程中极其重要。有研究发现,在结直肠癌细胞快速转移过程中,丝氨酸、甘氨酸和S-腺苷甲硫氨酸(SAM)的含量会出现增加的现象。这是由于结直肠癌细胞会通过诱导单泛素化修饰来增强PHGDH 的活性,间接促进结直肠癌细胞一碳单位代谢[47] 。与之类似的是,肿瘤细胞在SSP过程中,SSP 酶的表达或驱动丝氨酸合成增强的癌基因活化,会使细胞对丝氨酸和甘氨酸饥饿疗法产生抗性。因此,通过抑制PHGDH 的表达并协同丝氨酸和甘氨酸饥饿疗法,可以抑制肿瘤一碳代谢和癌症生长[48] 。同时,也有研究表明,SSP 过程中的磷酸丝氨酸转氨酶( phosphoserine aminotransferase,PAST)的缺失,或从饮食中去除丝氨酸对于抑制结直肠癌异种移植物的生长和增强5-氟尿嘧啶(5-fluorouracil,5-FU)的抗肿瘤活性极其重要[49] ,研究表明,内源性或外源性的丝氨酸都有助于结直肠癌的生长和肿瘤细胞对5-FU 的抗性。
肿瘤细胞内的甘氨酸不仅有多种合成方式,而且其合成途径中的许多代谢酶通过与转录因子相互作用参与肿瘤生长调控。例如,c-Myc 和活化转录因子4(activating transcription factor 4, ATF4)可以转录调控丝氨酸羟甲基转移酶(serine hydroxy methyltransferase, SHMT)影响肿瘤细胞生长[50] ,肿瘤抑制因子p53 也可以调控甲基四氢叶酸脱氢酶(methylenetetrahydrofolatedehydrogenase, MTFHD),以响应细胞增殖和DNA 损伤[51] 。值得注意的是,丝氨酸的缺乏不仅会导致甘氨酸合成受阻,还会影响肿瘤细胞的生长和核苷酸的合成。但是,与甘氨酸缺乏对肿瘤细胞的影响相比,线粒体甘氨酸的生物合成途径受阻会严重影响肿瘤细胞的增殖[52] 。此外,通过建立异种移植和同种异体移植的大鼠模型发现,大鼠摄取丝氨酸和甘氨酸的含量下降后,确实可以减少肿瘤细胞的生长[53-54] 。这一系列研究表明,丝氨酸或者甘氨酸的代谢对于肿瘤生长及其重要, 其关键代谢途径或代谢酶与肿瘤因子之间存在相互作用。因此,靶向丝氨酸或甘氨酸代谢或能成为肿瘤治疗的关键策略。
2. 4 精氨酸代谢
与正常人体细胞相比,某些具有营养缺陷型的癌细胞具有更高的营养需求。精氨酸作为一种多功能氨基酸,在代谢和信号通路中具有多种生物学功能,其在癌细胞中的含量和相关代谢通路对肿瘤细胞的生长具有重要影响。精氨酸是肿瘤细胞合成蛋白质合成所必需的,也是合成尿素、一氧化氮、多胺、脯氨酸和胍丁胺等几种分子的前体[55] 。有报道指出,精氨酸对于免疫系统应答、心血管以及肝肾系统正常功能等有重要作用[56] 。在正常细胞中,瓜氨酸经过一系列分解代谢形成精氨酸和琥珀酸,其中精氨琥珀酸合成酶(arginine succinate synthase, ASS)是精氨酸合成过程中的限速酶。然而,在大多数类型癌症中ASS 表达水平较低,呈现外源精氨酸依赖性,因而ASS 可作为治疗癌症的重要靶点[57]。研究发现,转录因子c-Myc 和缺氧诱导因子1α(hypoxiainducible factor-1α, HIF-1α)可以转录调控ASS1 的表达,并且抗癌药物顺铂能够通过上调HIF-1α 和下调c-Myc 来抑制ASS1 的表达,从而协同精氨酸饥饿疗法以抑制肿瘤细胞的生长[58] 。值得注意的是,最新研究发现,在ASS1 基因低表达的肝癌患者中,过表达ASS1基因能够通过激活蛋白激酶R 样内质网激酶(PKR-like ER kinase, PERK) / 真核起始因子2 的α 亚基(α subunit of eukaryotic initiation factor2, eIF2α) / ATF4 / C/ EBP 同源蛋白(C/ EBP-homologousprotein, CHOP)信号通路,抑制肿瘤生长并增强体外和体内药物化疗的疗效,且ASS1 的表达与患者良好的预后相关[59]。
有研究表明,许多实体瘤不能合成精氨酸,因而促进肿瘤细胞内精氨酸的降解为治疗癌症提供了另一思路。精氨酸降解后能够产生包括尿素、鸟氨酸和多胺等在内的代谢产物。其中多胺是小的有机聚阳离子,在DNA 复制、翻译和细胞增殖方面扮演着重要的角色[60] 。鸟氨酸脱羧酶1(ornithine decarboxylase1, ODC1)是多胺合成中的限速酶,影响肿瘤的发生发展,并在肿瘤细胞中出现高表达[61] 。值得注意的是,有研究发现,p53能够转录抑制氨甲酰基磷酸合成酶I ( carbamoyl phosphate synthetase,CPS Ⅰ)、鸟氨酸氨甲酰基转移酶(ornithine transcarbamylase,OTC)和I 型精氨酸酶(arginase I, ARGI)的表达,从而抑制肿瘤细胞尿素的产生和氨的解毒功能,导致细胞因氨毒性死亡[62] 。另外,ASS、ARGI和ARGII、诱导型一氧化氮合酶(iNOS)、精氨酸脱羧酶(arginine decarboxylase, ADC)以及过氧化氢酶(CAT)活性均显示能影响精氨酸的合成和分解代谢[63-64] 。研究表明,精氨酸代谢过程中的代谢物能够参与肿瘤细胞的尿素循环并受转录因子p53 的调控。
2. 5 半胱氨酸和胱氨酸代谢
半胱氨酸是一类半必需氨基酸,在正常细胞中,其不仅参与蛋白质合成,还会影响细胞的氧化还原稳态。然而,肿瘤细胞中的半胱氨酸和胱氨酸在为肿瘤细胞的增殖提供能量的同时,又会通过调控肿瘤细胞的信号传导介导肿瘤的生长。最近的研究表明,在某些情况下,半胱氨酸的缺乏不仅可以阻碍癌细胞的增殖,还可触发铁死亡[65] 。因而半胱氨酸饥饿疗法被用于抑制小鼠癌症模型中的肿瘤生长[66] 。在半胱氨酸代谢过程中,半胱氨酸双加氧酶已被证明与肿瘤的耐药性、不良预后相关[67-68] ;同时,其代谢过程中的胱硫醚β-合酶(cystathionine-beta-synthase,CBS)也被证明与恶性肿瘤的增殖、迁移和扩散有关[69-70] 。此外,在细胞培养基中去除半胱氨酸后发现极大地增强了单磷酸活化蛋白激酶(AMPactivatedprotein kinase, AMPK)活性;进一步研究发现,一种可直接结合半胱氨酸的半胱氨酰tRNA 合酶(cysteinyl-tRNA synthetase, CARS) 可以结合钙/钙调蛋白依赖性蛋白激酶激酶2(calcium/ calmodulindependent protein kinase kinase 2, CaMKK2),从而感知半胱氨酸的缺失并激活AMPK,最后调节肿瘤细胞生长[71] 。总而言之,这些研究表明,半胱氨酸及胱氨酸的代谢在肿瘤发生发展过程中具有调控功能。
2. 6 脯氨酸代谢
脯氨酸代谢在肿瘤代谢重编程中发挥重要作用,影响着癌症的发生和发展。脯氨酸代谢与肿瘤细胞中的ATP 产生、蛋白质和核苷酸合成以及氧化还原稳态有关。通过分析大量癌细胞系发现,脯氨酸饥饿或抑制脯氨酸生物合成酶会抑制细胞的克隆形成和致瘤的能力。同时,进一步研究发现,部分依赖外源脯氨酸的癌细胞表现出mTORC1 介导的4E结合蛋白1(4EBP1,一种细胞信号传导蛋白)信号轴的过度激活,以及内质网(endoplasmic reticulum,ER)应激反应;若外源补充脯氨酸可减轻ER应激,并促进细胞稳态和克隆形成[72] 。除此之外,在脯氨酸代谢酶中,吡咯琳-5-羧酸还原酶(pyrroline-5-car-boxylatereductase, P5CR) 在多种类型癌症中高表达,并且促进肿瘤的生长。而且,沉默调节蛋白3(sirtuin 3, SIRT3)在体内和体外均能与P5CR1 相互结合并去乙酰化P5CR1,增加P5CR1 的活性,从而促进肿瘤的生长[73] 。由于脯氨酸在癌症代谢中发挥特殊的作用,抑制脯氨酸的生物合成可以抑制肿瘤生长[74] 。
在肿瘤细胞中,脯氨酸的降解类似于其合成的逆反应,其中脯氨酸脱氢酶(proline dehydrogenase,PRODH) 是由肿瘤抑制因子p53 诱导的基因之一[75] 。脯氨酸分解代谢不仅支持3D 培养的肿瘤细胞生长,还能促进肿瘤细胞的转移;通过抑制脯氨酸代谢过程中的PRODH 可以抑制肿瘤的转移[76] 。而另有研究发现,在肿瘤细胞中过表达脯氨酸氧化酶(proline oxidase, POX)可以通过增加活性氧(reactiveoxygen species, ROS)表达诱导肿瘤细胞发生凋亡[77] ,这可能与脯氨酸代谢过程中肿瘤细胞的氧化还原状态相关。与之类似的是,肿瘤细胞在低氧条件下,POX 通过产生ROS介导自噬信号以维持肿瘤细胞的存活,进一步表明脯氨酸代谢是潜在的肿瘤治疗靶点[78] 。
2. 7 蛋氨酸代谢
在正常细胞中,蛋氨酸的摄取和代谢涉及包括甲基化反应、氧化还原维持、叶酸代谢和多胺合成在内的许多细胞生物学功能,这些功能能够协调细胞核苷酸的合成和氧化还原状态的稳定。在许多情况下,这些功能与癌症的发病机制相关。研究发现,蛋氨酸作为S-腺苷蛋氨酸( S-adenosyl methionine,SAM)的底物,是癌细胞重要的甲基供体和必需氨基酸, 肿瘤细胞在异常增殖过程中会大量摄取蛋氨酸,并代谢成为SAM 来维持其重编程的需要。研究发现,哺乳动物细胞需要活化的叶酸来产生用于生长和分裂的核苷酸。在癌细胞中,四氢叶酸(tetrahydrogenfolic acid, THF)能将一个碳单位转移到氨基酸、核苷酸或其他分子上,因此,在核酸和蛋氨酸的合成中具有重要作用。其中5-甲基四氢叶酸(5-methyl-THF)能够在钴胺素依赖性蛋氨酸合酶(methioninesynthase, MTR)的作用下,将同型半胱氨酸转换成蛋氨酸并生成THF。有研究发现,MTR 能够通过维持THF的代谢稳态,控制肿瘤细胞的生长[79-80] 。研究表明,MTR 不仅能参与蛋氨酸循环,还能通过调控蛋氨酸循环参与叶酸代谢,这为蛋氨酸的膳食干预与抗叶酸疗法的结合提供了思路。
2. 8 支链氨基酸的代谢
支链氨基酸( branched-chain amino acids,BCAAs)作为氮供体为肿瘤细胞的增殖需求提供核苷酸等大分子合成原料,对于癌细胞的生长是不可或缺的。BCAAs 除了直接参与蛋白质合成外,还能通过分解代谢产生谷氨酸等中间代谢物,从而与其他代谢途径相关联[81] 。例如,癌细胞中的BCAAs通过LAT1 进入细胞,然后由溶质载体家族25成员44(solute carrier family 25 member 44, SLC25A44)蛋白将其转运到线粒体[82] 。肿瘤细胞内的BCAAs 通过支链氨基酸转氨酶(branched-chain aminotransferase,BCATs)转化为包括α-酮异己酸(α-ketoisocaproate,KIC)、α-酮-β-甲基戊酸(α-keto-β-methylvalericacid,α-KMV)以及α-酮异戊酸(α-Ketoisovalericacid, KIV)在内的支链α-酮酸(branched-chainα-ketoacid, BCKAs),同时将氨基从BCAAs 转移到α-酮戊二酸(α-Ketoglutaric acid, α-KG) 生成谷氨酸。因此,BCATs 不仅能控制BCAAs 和BCKAs的水平,还能调控肿瘤细胞内α-KG 和谷氨酸的水平[83] 。肿瘤细胞中BCAAs 代谢的重编程是由参与BCAAs 代谢途径的BCAAs 转运体和代谢酶的表达以及活性的改变决定的。有研究报道称,BCAAs 代谢与胰腺导管腺癌(pancreatic ductal adenocarcinoma,PDAC)的发展有关,通过降低膳食中BCAAs 的水平或者采用支链氨基酸转氨酶2(branched chainaminotransferase 2, BCAT2) 的抑制剂,能减缓小鼠PDAC 的生长[84] 。此外,BCAT1 的启动子区含有多种转录调节因子的结合位点,包括c-Myc、HIF-1 和SMAD5[85-86] ,这些转录调节因子均可以通过调控BCAT1 进而调控支链氨基酸的代谢,并对肿瘤代谢产生影响。Fig. 1 为肿瘤细胞中的氨基酸代谢。
3 氨基酸代谢与肿瘤mTOR 信号通路
癌细胞中的mTOR 能够调节蛋白质翻译并参与肿瘤细胞的生长和自噬调控。因此,mTOR 信号的传递和激活对于肿瘤细胞的生长和代谢活动极其重要。值得注意的是,氨基酸是激活mTOR 所必需的,而mTOR 能通过调控物质的分解代谢过程来控制细胞生长。早期的研究发现,细胞培养基中的氨基酸缺失能够显著抑制mTOR 下游底物的磷酸化。研究表明,mTOR 能够感应氨基酸的变化[87] 。氨基酸作为蛋白质合成的关键原料,能够直接结合氨基酸受体被细胞感知,进而将信号传递给mTOR。因此,mTOR能通过响应氨基酸水平控制肿瘤细胞的生长[88] 。
3. 1 氨基酸转运体调节mTOR 活性
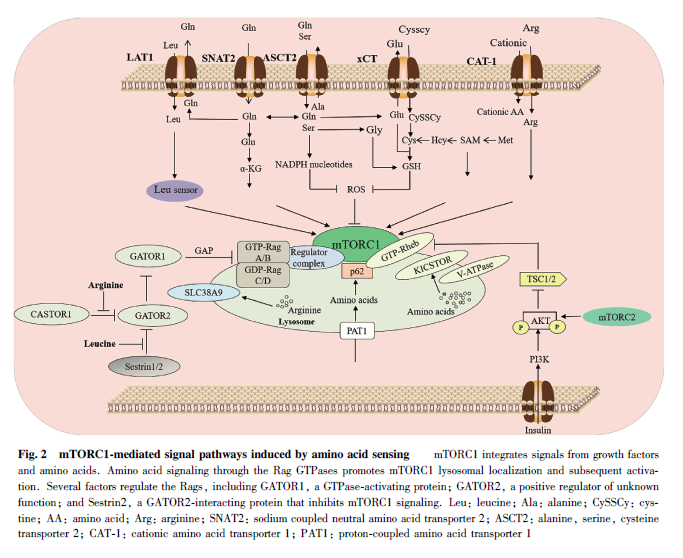
氨基酸转运体负责肿瘤细胞的内外氨基酸的交换,这意味着在肿瘤细胞在发生发展过程中,氨基酸转运体能够通过调节mTOR 活性控制肿瘤细胞的生长。研究发现,肿瘤细胞内mTOR 通过与其他分子形成复合物发挥生物效应,其包含两种复合体,分别为mTORC1 和mTORC2 复合物。肿瘤细胞内的多种氨基酸转运体在激活mTORC1 和促进细胞增殖过程中发挥关键作用(见Fig. 2)。例如,LAT1 通过激活mTORC1 的活性来促进体外和体内的癌细胞增殖,而LAT1 的敲除则会破坏肿瘤氨基酸的稳态,并降低mTORC1 的活性,从而减少小鼠异种移植肿瘤的生长[89] 。其他的研究也表明,LAT1 基因的敲除会导致骨肉瘤和胸腺瘤细胞发生凋亡[21, 90] 。这进一步证实了LAT1 对mTORC1 活性的调控功能。与之类似,在许多异种移植模型中也发现,ASCT2 基因的敲除会显著降低mTORC1 活性和肿瘤生长[91] 。除上述位于细胞质膜上的氨基酸转运体外,肿瘤细胞内的氨基酸转运体也能调控mTORC1 的活性。细胞内的质子偶联氨基酸转运蛋白1(proton-coupled amino acid transporter 1,PAT1)是位于溶酶体表面的氨基酸转运蛋白质,其会与RAS 相关的小G 蛋白( ras-related smallGTP- binding protein) 亚型 Rag GTP 酶发生相互作用,并在氨基酸依赖的mTORC1 激活中具有重要作用[92] 。最近的研究证实,PAT1 基因的过表达会活化mTORC1,并增强癌细胞对细胞周期蛋白依赖性激酶(cyclin-dependent kinases, CDK)4 /的耐药性[93] 。除此之外,溶质载体家族38 成员9 ( solute carrier family 38 member 9,SLC38A9)也被报道能在溶酶体膜上与Rag GTP酶相互作用,进而影响mTORC1的活性[94] 。这些研究均表明, 氨基酸转运体可以通过调控mTORC1 的活性影响肿瘤细胞的增殖。
3. 2 氨基酸调节mTOR 活性
近年来,除上述的氨基酸转运体外,细胞内氨基酸的精准感受器作为重要信号分子参与肿瘤细胞的代谢调控作用受到广泛关注。研究表明,GATOR2复合物是mTORC1 感知氨基酸信号通路中的关键正调控因子。亮氨酸的缺乏会通过亮氨酸传感器SAR1B 传递给mTORC1。在传递过程中,SAR1B 会通过自身构象的变化来靶向其激活剂GATOR2,从而间接调控mTORC1 活性,并在癌症的发展过程中发挥重要作用[95] 。与之类似的是,有研究发现,亮氨酸的缺乏会增强应激诱导蛋白Sestrin2 与GATOR2的相互作用,进而影响mTORC1 的表达[96] 。值得注意的是,在Sestrin2 与GATOR2 相互作用的过程中,泛素连接酶环指蛋白167(ring finger protein167, RNF167)和去泛素化酶STAM 结合蛋白样1(STAM binding protein-like 1, STAMBPL1)能协同调控Sestrin2 的多聚泛素化修饰水平,并影响Sestrin2调控mTOR 的机制,最后对结直肠癌的发生发展产生影响[97] 。其他的研究也逐步揭示,精氨酸、亮氨酸或者谷氨酰胺等均可以通过感应器或者代谢产生的ROS 对mTOR 产生作用,从而调控肿瘤细胞的增殖。然而,也有研究发现,与胱氨酸转运体SLC7A11介导的铁死亡不同的是,当肿瘤细胞中的半胱氨酸缺乏时,其可以通过抑制mTORC1 信号通路调控铁死亡[98] 。这进一步强调了氨基酸代谢对于mTOR活性调控的复杂性。这些研究表明,在肿瘤细胞代谢过程中,mTORC1 作为氨基酸的核心感应信号,在调节恶性肿瘤细胞的增殖和铁死亡等方面发挥了重要的生物学功能。Fig. 2 为mTORC1 介导的氨基酸感应的信号通路。
4 氨基酸代谢与肿瘤免疫
4. 1 氨基酸代谢与肿瘤微环境的关系
肿瘤的发生与发展,除了与自身细胞的恶性增殖相关外,还与其生存的环境有关,是影响肿瘤治疗的重要因素。其中,免疫细胞和免疫系统是肿瘤微环境的重要组成部分,一旦肿瘤微环境中的免疫细胞代谢发生改变或免疫系统受到破坏,就会导致肿瘤的恶化。研究发现,肿瘤细胞可以利用多种氨基酸来支持其增殖和转移。在这个过程中,肿瘤细胞中产生的下游代谢物被释放到肿瘤微环境中。一些代谢物表现出免疫调节特性,可以直接靶向和损害免疫细胞的功能。此外,转运蛋白质高表达肿瘤细胞会与免疫细胞竞争氨基酸,损害免疫细胞的功能[99-101] 。由于肿瘤细胞具备代谢重编程的能力,因此,能够适应肿瘤微环境(tumor microenvironment,TME)中的酸性或缺氧的不良环境。而包括NK 细胞在内的许多免疫细胞则需要葡萄糖来提供能量物质,以实现其抗肿瘤反应。然而,NK 细胞由于缺乏代谢适应性,在TME 中会受到代谢抑制。如果增加NK细胞在TME 中的代谢灵活性,将会提高NK 细胞的免疫毒性功能,并特异性增强抗肿瘤活性的机制[102] 。研究表明,TME 对于肿瘤细胞自身和免疫细胞极其重要。有趣的是,部分氨基酸及其代谢物既会影响肿瘤细胞的生长,又会对肿瘤微环境中的免疫细胞产生影响。例如,谷氨酰胺拮抗剂通过抑制肿瘤细胞的糖酵解代谢,在发挥抑制癌细胞生长功能的同时,又可以通过破坏肿瘤的免疫抑制微环境来克服肿瘤的免疫逃逸[103] 。与之类似的是,当调节性T 细胞受到谷氨酰胺的刺激时会增殖并上调IL-2 及IL-2 受体的表达,再通过一些中间代谢产物来促进骨髓来源的抑制性细胞(myeloid-derivedsuppressor cells, MDSCs) 成熟。最终,成熟的MDSCs会通过抑制肿瘤特异性T细胞的功能来介导肿瘤免疫逃逸[104] 。这意味着,谷氨酰胺不仅能够作为能量物质为肿瘤细胞和免疫细胞提供能量,而且,其在TME 中的含量会影响免疫细胞的成熟和免疫因子的释放。除谷氨酰胺外,TME 中的精氨酸和色氨酸代谢,也在肿瘤细胞的免疫调节方面扮演重要的角色。癌细胞和巨噬细胞会通过摄取和分解代谢,局部消耗TME 中的色氨酸,从而抑制抗原特异性T 细胞反应[105] 。与色氨酸一样,TME中细胞外精氨酸的可用性,会影响T 细胞抗肿瘤反应的幅度和持续时间[106] 。当癌细胞内的精氨酸被MDSCs 和巨噬细胞迅速分解代谢后,会表现出精氨酸缺乏的状态。而细胞毒性T 细胞对局部氨基酸可用性的波动很敏感,当局部精氨酸耗尽时,它们的效应物和抗肿瘤活性会显著降低[107-108] 。这些研究表明,TME 中的氨基酸代谢与肿瘤细胞的发生发展和肿瘤免疫密切相关。
4. 2 氨基酸代谢与免疫细胞功能的关系
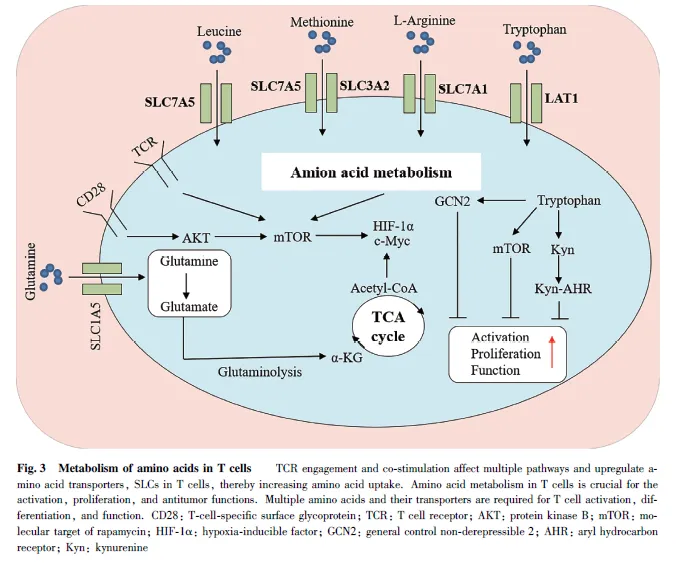
氨基酸代谢可以调控多种类型的免疫细胞功能,而且免疫细胞依赖于氨基酸代谢获得能量和生物基质,并通过激活其代谢过程中的关键酶或中间代谢物来维持其效应功能[109] 。由于细胞摄取氨基酸依赖于氨基酸转运体的精确调控,并且氨基酸转运体能控制免疫细胞功能,因此,不同类型的免疫细胞表达的转运蛋白质和受体存在差异,且不同类型的抗原和细胞因子会改变转运蛋白质的表达。例如,CD8+ T 细胞在细胞因子的作用下,通过T 细胞受体(T-cell receptor, TCR)的刺激并活化;由于活化的T 细胞需要更多的氨基酸供应来支持增殖,因此,活化的CD8+T 细胞表面溶质载体家族1 成员5(solute carrier family 1,member 5, SLC1A5)和溶质载体家族7 成员5(solute carrier family 7,member 5,SLC7A5)的表达量高于未活化的CD8+ T 细胞[110] ,而且氨基酸转运体的不对称分布也会影响T 细胞的分化[111] 。有趣的是,肿瘤细胞与免疫细胞在TME 中存在营养摄取竞争。当细胞通过mTORC1信号通路进行程序化的营养分配时[112] ,如果肿瘤细胞溶质载体家族43 成员2(solute carrier family43,member 2, SLC43A2)的表达水平升高,则会与T细胞竞争TME 中的蛋氨酸。一旦T 细胞缺乏甲硫氨酸,将会导致T 细胞的效应功能直接受损[113] 。与之类似的是,骨髓细胞是促进肿瘤生长和免疫逃避的肿瘤微环境的主要成分,通过抑制谷氨酰胺代谢能显著增加炎症性肿瘤相关巨噬细胞(TAM)的水平,并进一步导致肿瘤生长和转移减少[114] 。这进一步表明,氨基酸转运体对于免疫细胞功能的重要性。
最新的研究发现,氨基酸的供给对于免疫细胞的激活同样至关重要。天冬酰胺(asparagine, Asn)作为一种非必需氨基酸,能够通过直接结合淋巴细胞特异性酪氨酸蛋白激酶(lymphocyte-specific proteintyrosine kinase, LCK)来增强LCK 的信号传导和T 细胞的活化,并且无论在细胞水平还是大鼠体内均被证实,可以活化CD8+ T 细胞并增强抗肿瘤反应。反之,通过各种手段减少天冬酰胺的水平则会削弱CD8+T 的活性和效应T 细胞的产生[115] 。与天冬酰胺类似的是,无谷氨酰胺培养的T 细胞不能增殖或产生白介素2(Interleukin-2, IL-2)或γ-干扰素(Interferon γ, IFN-γ)[116] ,如果限制谷氨酰胺则会增加癌细胞中程序性细胞死亡受体-1(programmedcell death-1, PD-1)的水平,导致共培养的T 细胞失活[117] 。与之类似的是,丝氨酸能刺激谷胱甘肽(Lglutathione,GSH)合成,并维持调节性T 细胞(regulatoryT cells, Tregs)的功能[118] (见Fig. 3)。但与上述研究不同的是,在三阴性乳腺癌中,谷氨酰胺代谢基因与T细胞介导的细胞毒性标志物之间存在负相关,增加谷氨酰胺代谢可以降低T 细胞细胞毒性并提高肿瘤的存活率。因此,靶向肿瘤GLS 或减少谷氨酰胺代谢则可以改善抗肿瘤T 细胞的效应功能[119] 。除了氨基酸直接参与肿瘤免疫过程外,氨基酸代谢途径中的部分关键酶也会参与免疫细胞的功能调控。有研究发现,巨噬细胞在IFN-γ 或脂多糖(lipopolysaccharide,LPS)的刺激下,能促进信号转导和转录激活因子1(signal transducers and activator oftranscription 1, STAT1)介导的ASS1 转录,导致瓜氨酸消耗增多,从而介导炎症信号的激活。同时,瓜氨酸的增多会直接与非受体型酪氨酸蛋白激酶2(Januskinase 2, JAK2)结合,并抑制JAK2-STAT1 信号传导。研究表明,ASS1 在以消耗细胞瓜氨酸的方式控制炎性巨噬细胞活化和抗菌防御的过程中发挥关键核心作用,并通过其代谢物介导先天免疫信号的传递参与促炎反应的代谢检查[120] 。
5 靶向氨基酸代谢的肿瘤治疗
肿瘤细胞的代谢重编程是为了满足其增殖过程中的物质和能量需求。早期的癌症疗法使用抗代谢物来破坏肿瘤代谢,但随着对肿瘤代谢研究的深入,研究发现,许多癌症表现出偏好特定氨基酸的特征, 并且肿瘤细胞在增殖过程中,对外源氨基酸的供应极其敏感,通过减少血清中的氨基酸含量、限制氨基酸的转运或者通过分解氨基酸代谢酶的方式均能影响肿瘤细胞的生长。值得注意的是,与肿瘤细胞相比,正常细胞对氨基酸的需求较低,并且其在营养应激条件下合成目标氨基酸的能力较低。这使得靶向氨基酸代谢成为一种对正常细胞损伤小,且有效的手段[121] 。目前,氨基酸消耗疗法是通过使用异源、重组或工程化酶的方式来靶向氨基酸摄取和分解代谢,从而实现对肿瘤代谢重编程的调控。
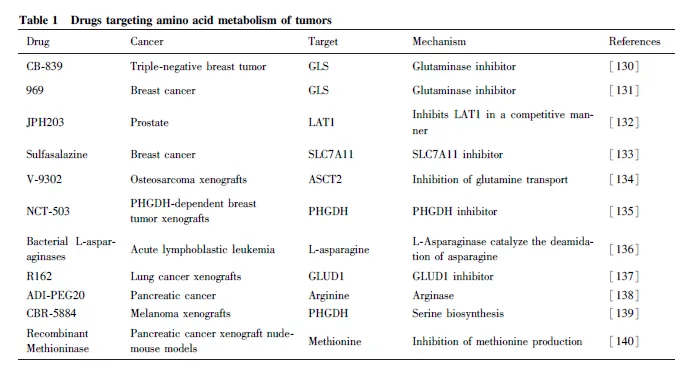
天冬酰胺(Asn)和谷氨酰胺对于肿瘤细胞的生长及其重要,其中Asn 饥饿对于诱导低水平表达或ASNS缺乏的白血病或淋巴瘤癌细胞作用明显[122] ,而ASNase单一疗法一直是治疗白血病和一些淋巴瘤病例的有效方法。有报道称,采用ASNase 与长春新碱、泼尼松和阿霉素可诱导细胞凋亡并治疗白血病[123-124] 。同时,鉴于Gln 在肿瘤细胞存活中的关键作用,已通过药理学GLS 抑制剂直接靶向该途径。在临床研究中,GLS 抑制剂CB-839 已被证明能够降低线粒体氧化磷酸化,抑制人急性髓系白血病细胞(acute myeloid leukemia, AML) 细胞的生长。癌细胞需要精氨酸来维持生长,因此,耗尽内源性Arg 是一种潜在的治疗干预措施。研究发现,使用聚乙二醇化形式的精氨酸脱亚胺酶(PEGylated argininedeiminase, ADI-PEG20)或精氨酸酶靶向Arg 代谢,或尿素循环的两种关键酶来治疗肿瘤是一种有效的策略[125] 。同时,ADI 将Arg 降解为瓜氨酸在预防黑色素瘤、肺癌、肾细胞癌和肝细胞癌等肿瘤细胞增殖方面具有有益作用[126-127] 。除此之外,Cramer等设计了与PEG 偶联的半胱氨酸酶,可降低血清中的半胱氨酸水平并消耗细胞外半胱氨酸,逐渐成为一种癌症的治疗工具。研究发现,半胱氨酸酶处理肿瘤细胞后可通过抑制mTOR 活性,并介导细胞内ROS 积累增加和自噬增加, 从而发挥抑癌功能[128-129] 。这一系列的研究表明,在癌症的治疗过程中,通过设计靶向肿瘤氨基酸代谢过程中的转运体、代谢物或代谢酶已成为一种有效的治疗手段。Table 1 为靶向肿瘤氨基酸代谢的药物 。
6 问题与展望
肿瘤细胞在异常增殖过程中,为获取更多的物质和能量,通常需要通过改变自身的代谢方式或增加代谢量。其中,肿瘤代谢重编程不仅涉及到葡萄糖和脂质代谢,还涉及到氨基酸代谢的改变。在肿瘤细胞中,氨基酸一方面能直接作为蛋白质合成的成分,另一方面又能作为能量代谢的中间产物参与肿瘤细胞的生长过程。在调控肿瘤功能方面,不仅可以通过细胞内的氨基酸转运体、感应器或者自身代谢产生的中间产物、酶等参与许多生物学过程,还能通过调控代谢途径改变肿瘤微环境或细胞内信号通路,从而实现对肿瘤细胞的生长调控作用。值得一提的是,氨基酸的代谢对于免疫细胞的激活或者分化等具有重要的生物学功能,可以通过调控免疫细胞及免疫因子的产生对肿瘤免疫产生影响。这不仅体现了氨基酸作为药物治疗靶点在治疗肿瘤方面的巨大潜力,也进一步表明可以通过调控氨基酸代谢为肿瘤免疫提供治疗辅助。
随着研究的深入,发现相较于正常细胞来说,肿瘤细胞对于氨基酸的需求和依赖性更加偏重。因此,针对氨基酸代谢治疗肿瘤的方式更加有效,且对正常细胞的损伤较小。目前,临床研究已开发出多种针对氨基酸转运体、谷氨酰胺、精氨酸等在内的氨基酸或氨基酸代谢酶的试剂,部分药物对肿瘤细胞表现出良好的疗效。然而,由于部分针对氨基酸代谢酶的天然药物存在半衰期短、疗效差与药物抗性等问题,设计针对氨基酸代谢的有效药物仍存在问题。值得注意的是,肿瘤细胞的发生与发展通常由多种因素相互作用所致,而氨基酸及其代谢物参与调控的肿瘤诱导因素包括肿瘤微环境中的信号分子的传导,肿瘤代谢重编程的发生,持续增殖能力的获得以及免疫反应的产生。因此,未来研究肿瘤中的氨基酸代谢将更应该关注氨基酸代谢的关键酶、氨基酸代谢的多种信号通路,及其与葡萄糖、脂质代谢的关联,为寻找肿瘤细胞内氨基酸代谢的感应、反馈机制及药物靶点奠定理论基础。
 电话:021-60838199
电话:021-60838199 客户服务专线:400-839-8199
客户服务专线:400-839-8199 传真:021-60705089
传真:021-60705089 地址:上海市闵行区三鲁公路3279号
地址:上海市闵行区三鲁公路3279号